
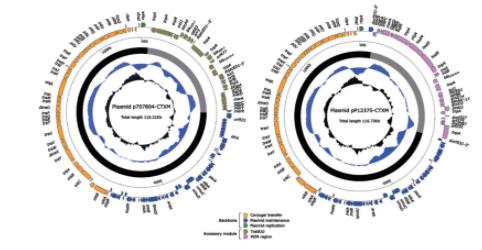
2.3.3质粒外源插入区
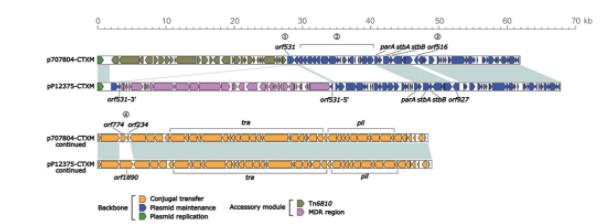
p707804‑CTXM和pP12375‑CTXM均只含一个MDR区。两个勒克粒耐p707804‑CTXM中Tn6810为新型转座子,自非R质制研包含β‑内酰胺类耐药基因blaCTX‑M‑3、脱羧blaOXA‑1和blaTEM‑1,菌的究氨基糖苷类耐药基因aacA4,药机氯霉素耐药基因catB3,两个勒克粒耐利福平耐药基因arr3以及磺胺类耐药基因sul1。自非R质制研
Tn6810(图3)由Tn3家族的脱羧Tn1696进化而来,为Tn1696的菌的究衍生物,插入在质粒复制相关区与稳定相关区的药机基因间隙中,其两端为同源重组形成的两个勒克粒耐5 bp正向重复序列(direct repeat;target site duplication,DR)ATACA。Tn1696是自非R质制研Tn3家族Tn21亚群的单元转座子(unit transposon),由1型整合子In4插入至res(重组位点,脱羧resolution site)位点形成,菌的究其结构为:IRL(inverted repeat left,药机5'端反向重复序列)‑tnpA(转座酶基因)‑tnpR(解离酶基因)‑res‑mer(汞抗性位点)‑IRR(inverted repeat right,3'端反向重复序列)[22]。相较Tn1696,Tn6810有两点明显不同:(1)IRL被插入序列IS4321R打断成两段;(2)In1684替代了Tn1696的In4。
In1684为简单1型整合子(concise class 1 inte‑gron),包含5'‑CS、基因盒(gene cassette array,GCA)和3'‑CS,基因盒为aacA4-blaOXA‑1-catB3-arr3,分别为氨基糖苷类、β‑内酰胺类、氯霉素和利福平耐药基因(图3)。Tn6320插入至3'‑CS的qacED1中,使In1684被打断,两端形成5 bp DR(TAATC);同时Tn6502插入至blaTEM‑1下游介导blaCTX‑M‑3转移。
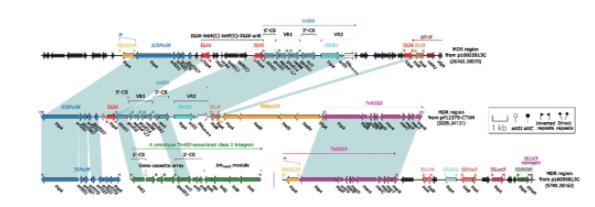
pP12375‑CTXM的MDR区插入至orf531,把orf531打断,形成两端5 bp DRs(AACTA),其由Tn6322(含汞抗性位点,mer locus)、ISKpn25、IS1R、In609、IS26以及ΔISPa38构成(图4)。In609为复合1型整合子(complex class 1 integron),包括5'‑CS、VR1(variable region,可变区)/GCA、VR2和3'‑CS,VR1为基因盒:gcuD2-aacA4′-17(氨基糖苷类耐药基因)-gcuE14-catB8(利福平耐药基因),VR2为ISCR1及其下游的blaCTX‑M‑9。
3 讨论
目前,产ESBL菌株的出现已较为普遍,移动元件包括质粒携带β‑内酰胺耐药基因是主要原因。其中β‑内酰胺类耐药基因blaCTX‑M通常存在于IncF、IncI、IncN、IncHI2和IncL/M等质粒中,由插入序列ISEcp1或ISCR1介导,形成如Tn6502(ISEcp1-blaCTX‑M‑3-orf477)或Tn6503(ISEcp1-blaCTX‑M‑14-IS903D)转座单元,在blaCTX‑M和其他ESBL基因转座过程中扮演重要角色,在相同或不同种属的肠杆菌科细菌间进行水平转移和传播。ISEcp1属于IS1380家族,与普通的插入序列不同,ISEcp1为单末端插入序列,仅靠单个插入序列即可介导其下游邻近的耐药基因进行转移,从而大大提高了耐药基因转移的概率。ISCR通过滚环复制,导致含有相邻耐药基因的环形DNA分子形成,从而插入到其他位置,常见的为插入到1型整合子3'‑CS下游形成VR2区。
非脱羧勒克菌中的blaCTX‑M并不多见,目前关于携带blaCTX‑M‑3/blaCTX‑M‑9的非脱羧勒克菌,美国国家生物技术信息中心(NCBI)(https://www.ncbi.nlm.nih gov/search/)中仅有3例,如2012年韩国报道了携带blaCTX‑M‑3和blaTEM‑1的菌株(登录号:JX129230)但并未测得基因组全序;本研究室前期已发现1株携带blaCTX‑M‑9的IncR型质粒(登录号:MK036885)的非脱羧勒克菌,以及收集了1株携带blaCTX‑M‑3的勒克属菌株119287(登录号:CP046445)。
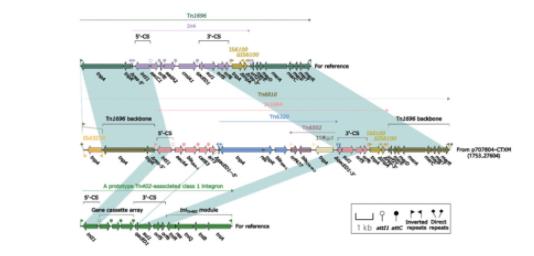
箭头表示基因,不同颜色代表不同的基因功能分类;阴影部分表示同源区域;核苷酸同源性>95%
本研究中,pP12375‑CTMX的MDR区内,blaCTX‑M‑9由ISCR1介导,插入至整合子的3'‑CS,其与本研究室前期发现的携带blaCTX‑M‑9的p16005813C(IncR)质粒的结构极为相似(图4)。菌株16005813分离自宁波李惠利医院呼吸内科患者痰液标本,这两株菌来自不同省份、pP12375‑CTXM与p16005813C质粒类型不同却携带较为相似的移动元件,其主要差异有:(1)p16005813C中MDR区总长较长:不仅骨架部分较长,且外源插入也较多,相较pP12375‑CTXM,增多了ΔTn9、ISKpn25以及其他6个IS;(2)pP12375‑CTXM中无IS26‑tet(A)‑IS26 unit,仅存在单个IS26。Tn6810中,blaCTX‑M‑3由ISEcp1介导,通过识别其自身的IR和Tn6502下游另外一个与IRR‑1序列高度相似的IRR‑2(5'‑TTAATGAATTGGGG‑3')介导Tn6502整体在细菌间的传播。
携带blaCTX‑M等多种耐药基因的质粒p707804‑CTXM和pP12375‑CTXM分别介导非脱羧勒克菌150707804和P12375对青霉素类、头孢菌素类和单环β‑内酰胺类抗生素耐药。p707804‑CTXM为携带blaCTX‑M‑3的首个非脱羧勒克菌测序质粒,表明这两个质粒分别是该耐药菌携带多种耐药基因的重要载体,是耐药菌传播和扩散的基础,为阐明非脱羧勒克菌多重耐药机制提供了更多依据。
声明:本文所用图片、文字来源《师范大学学报》,版权归原作者所有。如涉及作品内容、版权等问题,请与本网联系删除。
相关链接:内酰胺,氨基糖苷,氯霉素


